Document Type : Original article
Introduction
Genistein is the soy isoflavone1 which can pass across the blood-brain barrier like estradiol2. Several studies reported that genistein plays a role as a powerful neuroprotective in central nervous system3-6. Also, it could bind to estrogen receptors and has been reported to act like an agonist of both α and β estrogen receptors from different studies on cell culture, estrogen receptor, reporter-genes and animal models7. In our previous experiment, genistein was able to significantly increase the seizure threshold induced by pentylenetetrazole in Ovariectomized (OVX) mice. This anticonvulsant impact of genistein was prohibited by selective antagonist of estrogen receptor8. Also, genistein treatment has neuroprotective effects on status epilepticus-induced hippocampal damage in OVX female rats and preserved the inhibition of the dentate gyrus2,7. Menze et al in 2015 revealed a neuroprotective and memory enhancing effects of genistein in a rat model of Huntington’s disease. Also they suggest these impacts of genistein was mediated by its antioxidant, anti-inflammatory and cholinesterase inhibitory activities3. Moreover, it was shown that genistein inhibited glutamate-induced apoptosis in mouse primary neuronal cell cultures4.
Genistein is known as a powerful phytoestrogen. Phytoestrogens can obviously increase the expression of Nitric Oxide Synthase (NOS) in hippocampus playing a protecting role against neurodegenerative disease9. Pretreatment with genistein and 17β-estradiol in Alzheimer’s disease decreased the expression of inducible Nitric Oxide Synthase (iNOS) in cultured astrocytes10. Furthermore, our previous study has shown nonselective nitric oxide synthase inhibitor blocked the anticonvulsant activity of oleuropein, which is another well-known phytoestrogen11.
In addition, N-methyl-D-aspartate (NMDA) receptors have received very much attention because of their involvement in neuronal development, neurodegenerative diseases, and neuronal excitotoxicity12-14. Genistein significantly inhibited glutamate-induced apoptosis and the effect of this isoflavone was most prominent in the hippocampal cells5. Earlier studies revealed that genistein not only significantly depressed ischemia/reperfusion-related release of glutamate among rat’s cerebral cortex, but also reduced the influx of Ca after cerebral ischemia through inhibiting tyrosine-phosphorylation of NMDA receptors15.
In present study, we intended to assess the possible neuro-protective mechanisms of genistein on the seizures threshold through NMDA receptors/NO pathway.
Materials and Methods
Animals
Mice (23-30 g) were obtained from Department of Pharmacology of Tehran University of Medical Sciences. Animals were accommodated under standard laboratory conditions including temperature (23±1◦C), a 12-hr-dark/12-hr-light cycle in polycarbonate cages with free access to standard animal chow and water. The study was conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication NO. 85-23; revised 1985).
Materials
The following materials were used: Genistein, pentylenetetrazole (PTZ), MK-801(an uncompetitive antagonist of the N-Methyl-D-aspartate (NMDA) receptor, a glutamate receptor), L-NG-Nitro-L-arginine
methyl ester hydrochloride (L-NAME, a non-selective inhibitor of NOS), L-arginine (a NO precursor) that all were purchased from Sigma (St. Louis, MO, USA). Ketamine hydrochloride (a nonbarbiturate anesthetic) was obtained from Rotexmedica (Trittau, Germany) and xylazine hydrochloride purchased from Alfasan (Woerden, The Netherlands).
The chemicals (L-NAME, L-arginine and ketamine) were dissolved/diluted in sterile isotonic saline solution; Genistein and Mk-01 dissolved in dimethyl sulfoxide (DMSO)/PBS at a ratio of 1:6. All chemicals were administered intraperitoneally (i.p.) in a volume of 10 ml/kg for the mice body weight, however pentylenetetrazole (PTZ; 0.5%) was excepted and injected intravenously (i.v.). Appropriate vehicle controls were prepared for each experiment. At least eight animals were used for each treatment group and each mouse was tested only once.
All behavioral tests were performed after a recovery period of 14 days of induction for ovariectomized16-18.
Determination of seizure susceptibility
A more sensitive method of i.v. administration of PTZ that allows better detection modulatory effects on convulsive tendency was used to assess the seizure susceptibility among mice. The threshold of PTZ was determined by inserting a 30-gauge butterfly needle into the tail vein of mice while they were restrained in a mouse restrainer. The needle was then secured to the tail with a narrow piece of adhesive tape, with the mouse moving freely. PTZ (0.5%) was infused at the constant rate of 1 ml/min using an infusion pump. Infusion was halted when forelimb clonus followed by a full clonus of the whole body was observed and the dose of PTZ administered (mg/kg of mice weight) was measured as an index of clonic seizure threshold knowing that the seizure threshold is dependent on PTZ dosage and is also time-related19.
Experiments
Intraperitoneal (i.p.) administration of genistein (10 mg/kg) was performed in three different time intervals (15, 30, 45 min). Then different doses of genistein (5, 10, 30 mg/kg, i.p.) was injected 30 min prior to induction of seizure. Doses and time points were determined based on a pilot experiment. Control animals received DMSO 1%. Administration of L-NAME (10 mg/kg, i.p.) and 7-nitroindazole (25 mg/kg, i.p.) was carried out 15 min before sub-effective dose of genistein (5 mg/kg). L-arginine (60 mg/kg, i.p.) was injected to mice 15 min before vehicle and genistein (10 mg/kg) groups. MK-801 0.05 mg/kg (i.p.) and ketamine 0.5 mg/kg (i.p.) were administered in mice 15 min prior to injection of vehicle- and genistein- treated mice. In the last experiment, diazepam (0.05 mg/kg, i.p.) was used as a positive control.
Statistical analysis
Data are presented as mean ± SEM of PTZ minimal dose (mg/kg) and analyzed using the SPSS statistical software package (Version 22). The data of PTZ-induced seizure thresholds were analyzed by one or two way analysis of variance (ANOVAs) followed by post hoc Tukey’s tests in two group comparison and multiple group comparison, respectively. The tests of homogeneity of variance were used to ensure normal distribution of the data. A p-value less than 0.05 were defined as statistically significant level.
Results
The effect of time and different doses of genistein on the seizure threshold
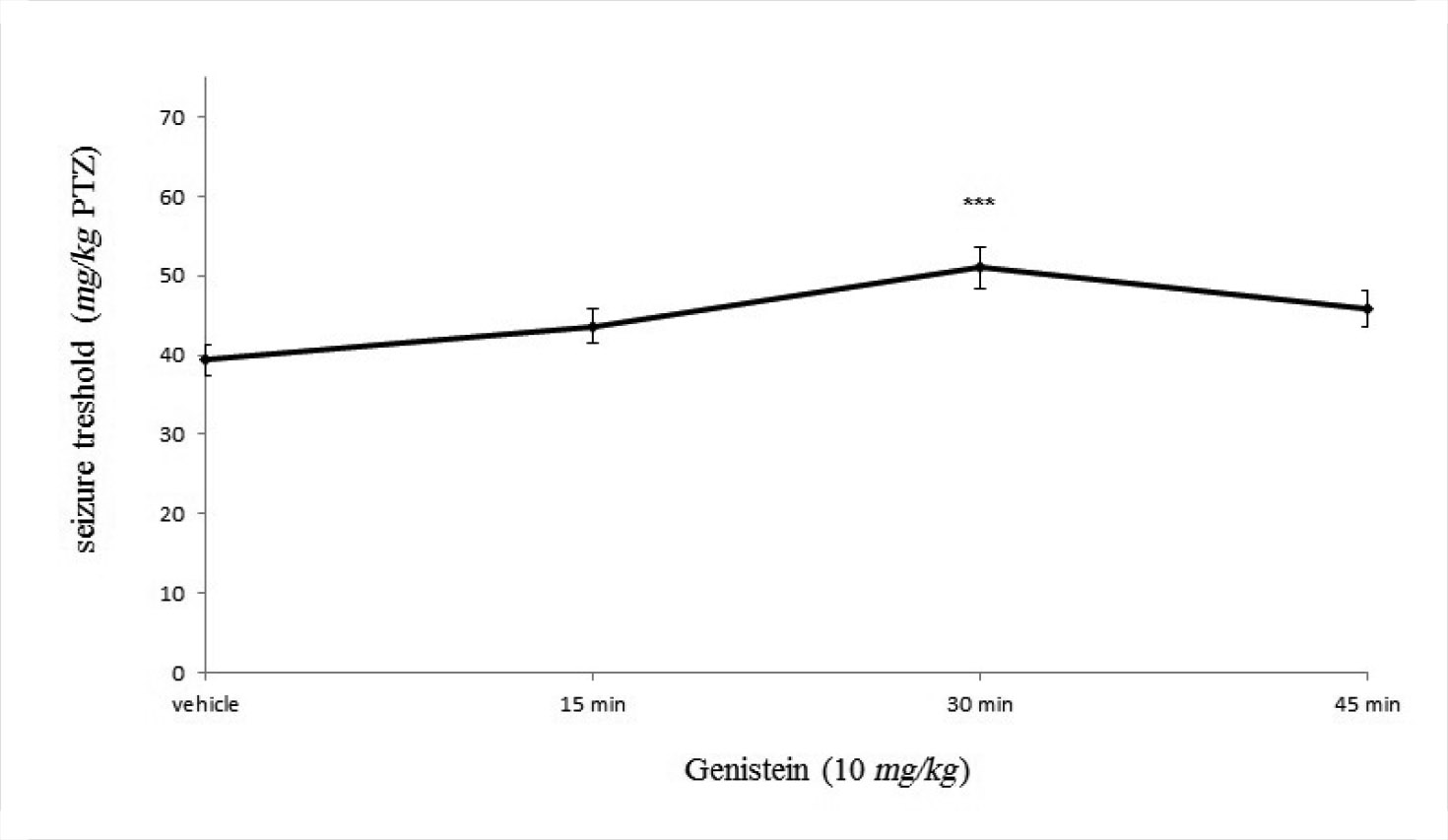
Figure 1 shows the time of the genistein effect on the seizure threshold. As shown, intraperitoneal administration of genistein (10 mg/kg) did not affect the PTZ-induced seizure threshold at 15 and 45 min, whereas it significantly reduced the seizure threshold 30 min after administration (p<0.001).
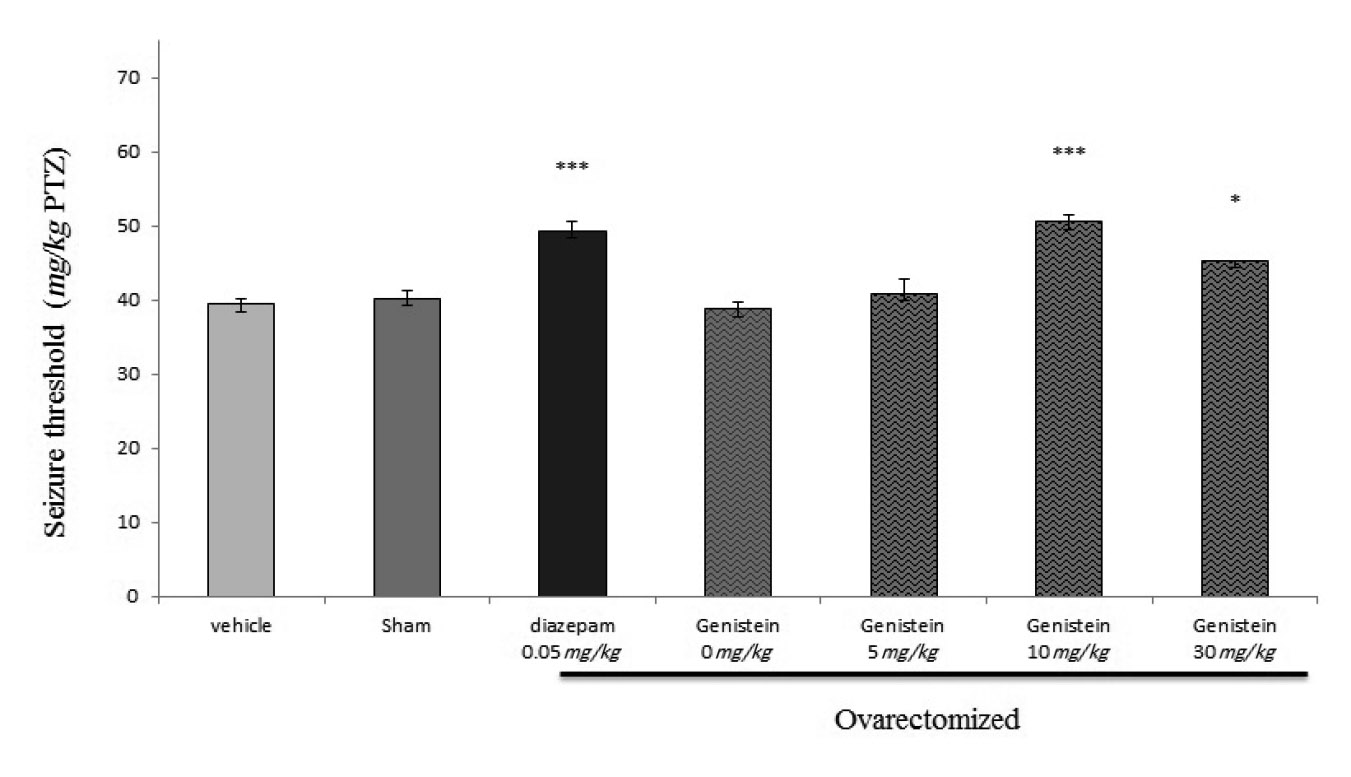
Figure 2 illustrates the effect of acute intraperitoneal administration of genistein (5, 10, and 30 mg/kg, i.p.) on the seizure threshold, when it was administered 30 min prior to PTZ. Genistein (10 mg/kg) significantly increased the PTZ-induced seizure threshold compared with vehicle-treated control animals (p<0.001).
Impacts of nitric oxide pathway on the anticonvulsive effect of genistein in PTZ-induced seizures in mice
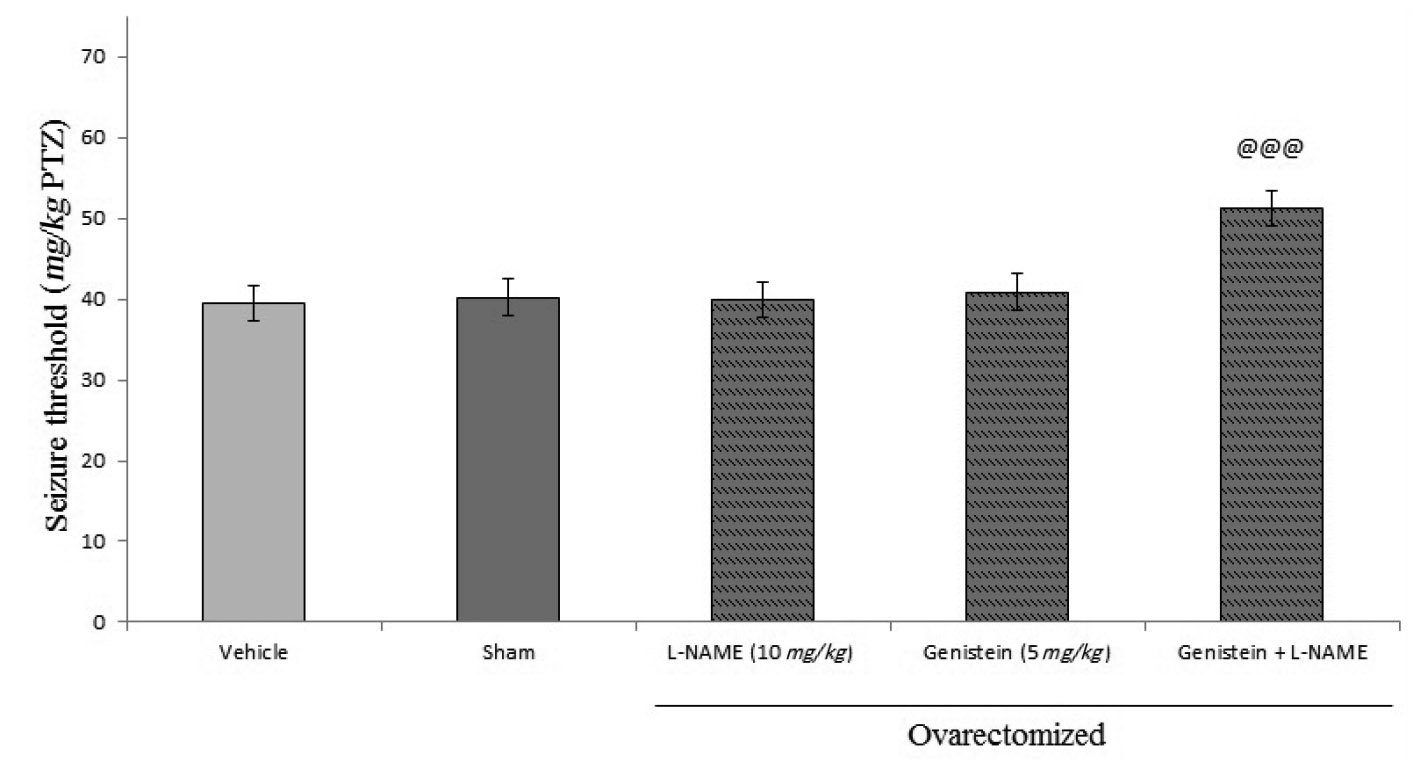
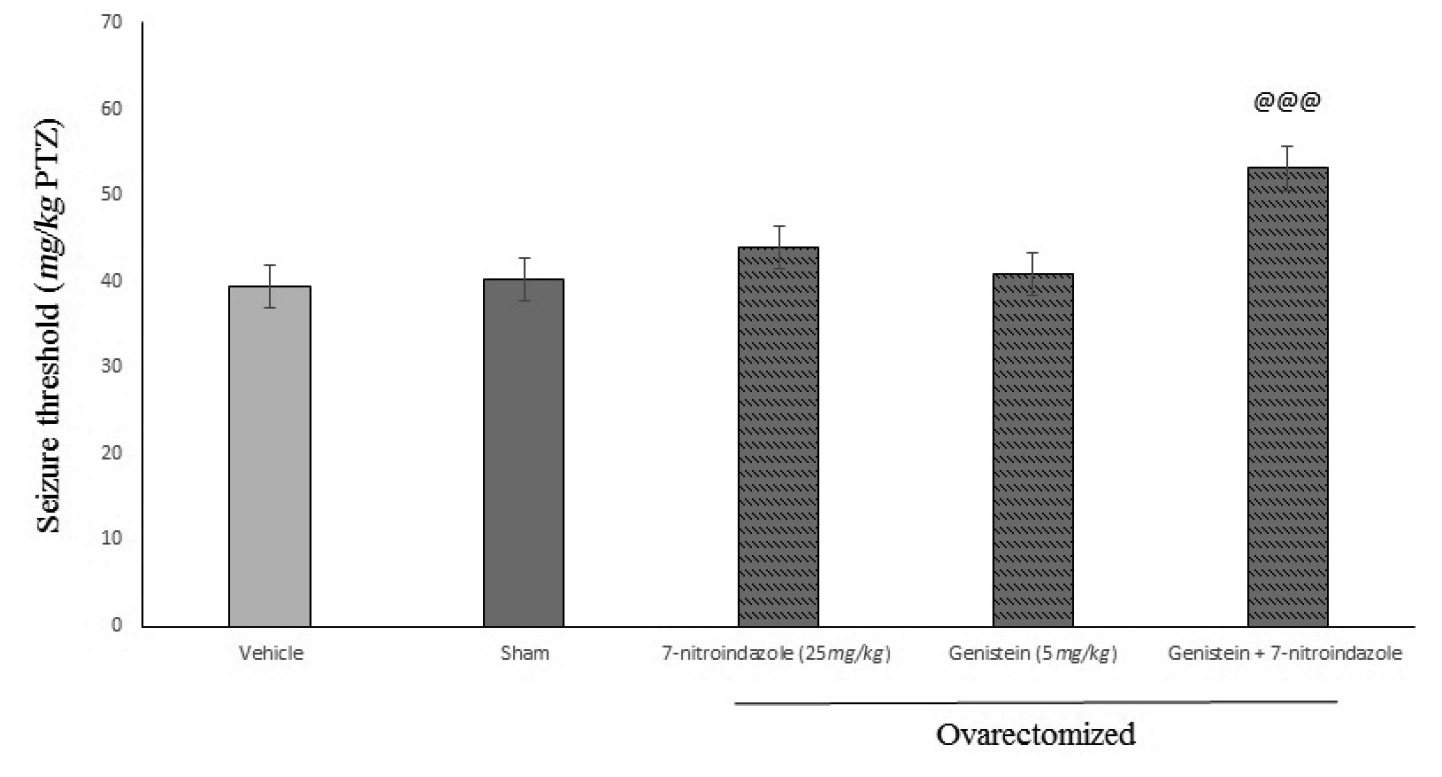
Figures 3 and 4 shows the effects of sub-effective dose of L-NAME and 7-nitroindazole pretreatment on sub-effective dose of genistein on seizure threshold in PTZ-induced seizures in mice. Administration of L-NAME (10 mg/kg, i.p.) and 7-nitroindazole (25 mg/kg, i.p.) by their own had no effect on seizure threshold (p>0.05) in comparison with vehicle treated controls. While, administration of both of them 15 min before sub-effect dose of genistein (5 mg/kg, i.p.) significantly increased the anti-convulsant effect of genistein (p<0.001).
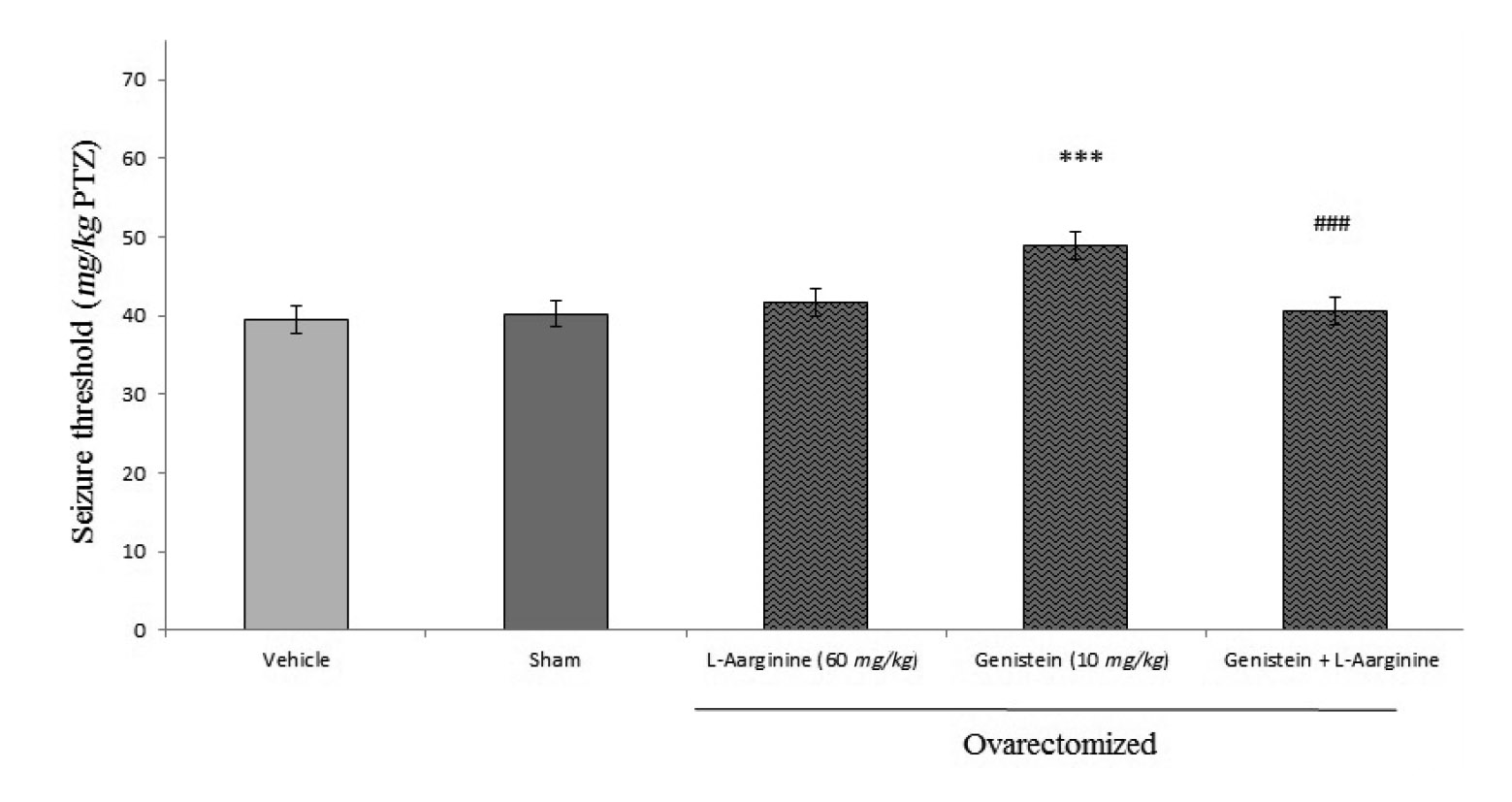
Figure 5 shows the L-arginine pretreatment effect on the anticonvulsant activity of genistein against PTZ-induced seizures. As shown in this figure, L-arginine pretreatment significantly reversed the anticonvulsant effect of genistein (p<0.001). Administration of L-arginine by its own had no effect on seizure threshold in comparison with saline treated controls.
Impacts of N-methyl-D-aspartate (NMDA) receptors on the convulsive effect of genistein in PTZ-induced seizures in mice
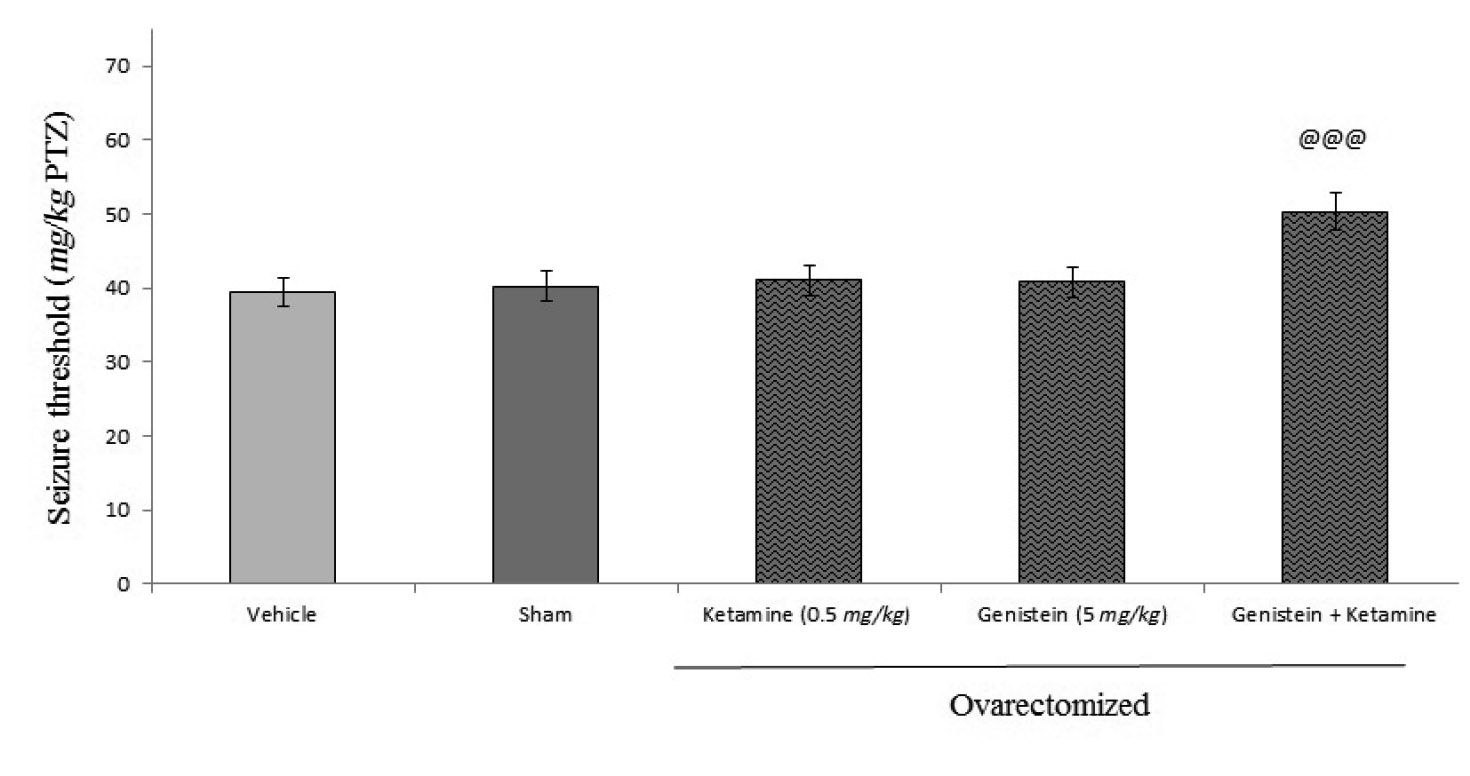
As shown in figure 6, administration of ketamine as a non-competitive antagonist of NMDA receptor (0.5 mg/kg, i.p.) did not alter the seizure threshold induced by PTZ in mice, while acute administration of ketamine (0.5 mg/kg, i.p.) 15 min before genistein (5 mg/kg, i.p.) significantly enhanced the anticonvulsant effect of genistein ((p<0.001).
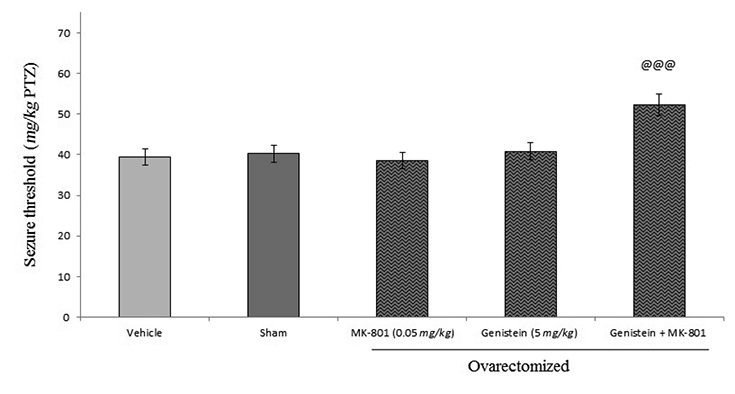
Furthermore, acute injection of MK-801 (a potential noncompetitive antagonist of NMDA receptor) 15 min before genistein (10 mg/kg, i.p.) significantly increased the anticonvulsant effect of genistein (5 mg/kg, i.p.) (p<0.001; Figure 7), whereas, MK-801 by its own had no significant effect on seizure threshold.
Discussion
We found that genistein have anticonvulsant characteristics in PTZ-induced seizure model in ovariectomized mice. L-arginine inhibited genistein effect on seizures threshold while L-NAME and 7-nitroindazole with sub-effective dose of genistein exacerbated the anticonvulsant effect of sub-effective dose of genistein.In our study, blocking of NMDA receptor with either ketamine or MK-80 led to inhibition of anticonvulsant effect of genistein. We, then, propose that genistein can modulate the anticonvulsant effect throughout NMDA receptors and nitrergic pathway in ovariectomized mice.
Recently Westmark et al investigated the effect of soy-diet on seizure in neurologic disorders such as Alzheimer’s disease, Down syndrome and Fragile X Syndrome (FXS) in audiogenic seizure mouse models. Surprisingly they found that daidzein exacerbated seizure events in mice which suffered from neurologic disorders20. In our previous study, we found that estrogenic receptors are involved in genistein anticonvulsant effect among mice after ovariectomy8.
Nitric oxide has been reported to act as a neuromodulator that exhibits dual pro-convulsive19,21 and anticonvulsive11,22 properties in PTZ model. In the present study we found that L-arginine inhibited genistein effect on seizure while L-NAME with sub-effective dose of genistein exacerbated the anticonvulsant effect of non-effective dose of genistein. Our results show that NO modulation is involved in the anticonvulsant effect of genistein so these results come in line with previous studies that have shown that nitric oxide system modulation was one of the most used mechanisms that genistein delivers its action through them5,12,23. Valsecchi et al concluded that NOS inhibition could be one of the multiple mechanisms that genistein relieved neuropathic pain, with both peripheral and CNS origins23. Liu et al demonstrated that genistein had direct non genomic effects on NOS activity on vascular endothelial cells leading to NOS activation that these effects were mediated by protein kinase activity and were unrelated to an estrogenic effect3. Gingerich et al revealed that genistein decreased the number of nNOS-positive neurons in PVN through activation of beta estrogen receptors by genistein24. Estrogen has an effect on NMDA in brain25. NMDA/NO cascade is a well-known pathway in brain where NMDA mediates seizure events and behavioral changes by activating NO system26-29.
In our study, blocking NMDA receptor by either ketamine or MK-801 led to inhibition of the anticonvulsant effect of genistein. These findings come in line with previous studies that showed NMDA receptor was implicated in mediating some of genistein effects. It has found that genistein inhibited NMDA-dependent cGMP production with high potencies30-32. Another study concluded estrogen receptor-mediated stimulation of the nNOS/PSD-95/NMDA receptor was likely to be a critical component of the signaling process by which estradiol facilitated coupling of glutamatergic fluxes for NO production in neurons33. Taken together, to explain our findings we suggest that mechanisms which underlines the anticonvulsant effect of genistein at least is partly carried out by modulating estrogen receptor/NMDA/ NO cGMP pathway which at the end leads to regulation of NO synthesis in brain.
Conclusion
In conclusion, the current findings underline that genistein had anticonvulsant activity in PTZ-induced seizures. This phenomenon could be modulated by NMDA receptor antagonists and nitric oxide synthase mediators which suggests the involvement of NMDA/ nitrergic pathways within the anticonvulsive impacts of genistein in the PTZ model of clonic seizures among ovariectomized mice.
Acknowledgments
This study was supported by a grant from Experimental Medicine Research Center, Tehran University of Medical Sciences (grant No. 91-02-158-18207).
Conflict of Interest
Authors declare that there is no conflict of interest.